
Project C2
Doctoral researcher: Sarah Gürtler
Principle investigator: Uwe Lendeckel
Co-supervisor: C. A. Helm
The AngIV-IRAP/AT4R alternative RAS axis - Implications for mitochondrial and beta-cell function
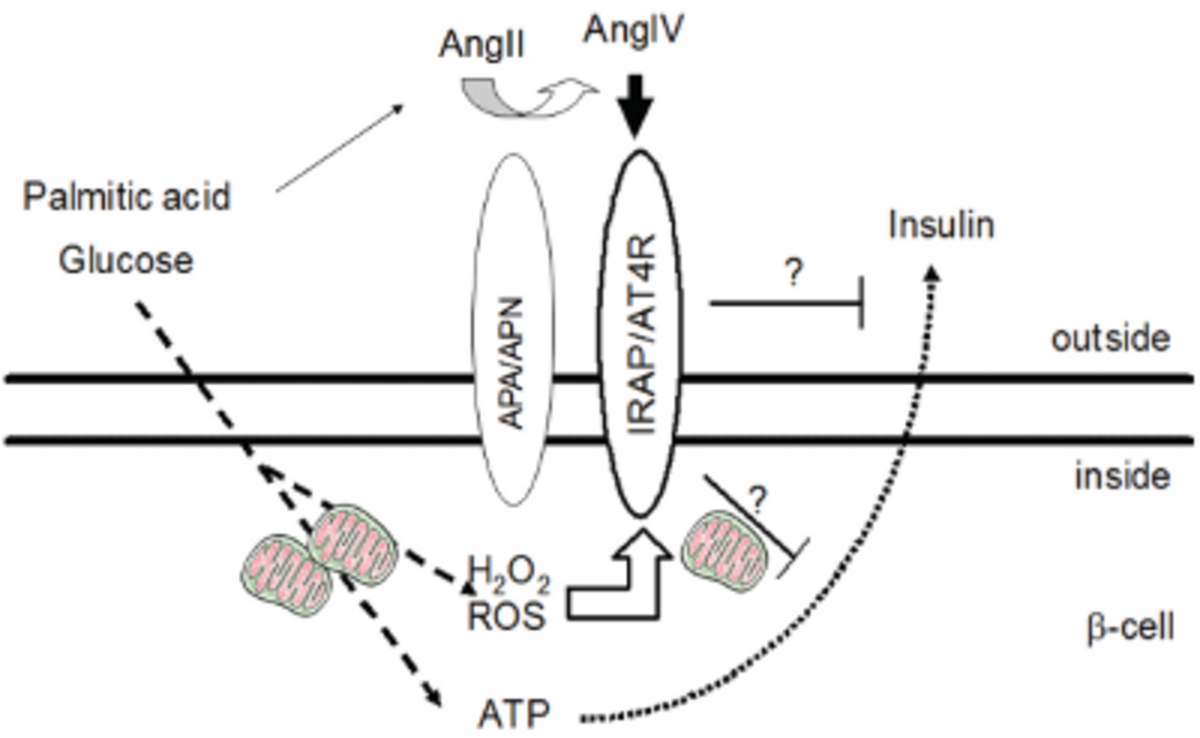
Hyperglycemia, hypertension and obesity, major components of the metabolic syndrome, are all associated with increased systemic and tissue levels of angiotensin II (AngII), the main effector peptide of the classical renin-angiotensin-system (RAS). Whereas it is well established that Ang II, via binding to AT1R, leads to increased production of superoxide anions and impairs glucose-stimulated insulin secretion (GSIS) and insulin signalling, the contribution of alternative RAS-axes to beta cell function remains to be elucidated fully. We have previously shown that BRIN-BD11 rat insulinoma cells and isolated mouse islets express all essential components of the AngIV-IRAP/AT4R alternative RAS axis, including aminopeptidases (AP) A and N, as well as insulin-regulated AP (IRAP) [1]. Exposure of BRIN-BD11 cells to high glucose increased cardiolipin oxidation and cytochrome c release in parallel to compromising mitochondrial membrane fluidity [2]. Notably, high glucose – besides inducing insulin production and release - dose-dependently increased mRNA and protein expression of IRAP, also functioning as receptor for AngIV, AT4R. Expression/activity of APA and APN, which together catalyze the conversion of AngII to AngIV and, thereby, generate the ligand for IRAP/AT4R, were equally elevated in response to high glucose. IRAP/AT4R is trans-located from intracellular GLUT4-vesicles to the surface in response to insulin, a process reported to be compromised in diabetes type 2. The role of IRAP/AT4R for beta cell function is unclear, as is that of the AngIV-IRAP/AT4R RAS axis in general and of APA and APN in particular.
This interdisciplinary project aims to elucidate the role of this RAS axis for beta cell function by studying the effects on ROS production, CL-oxidation, mitochondrial membrane fluidity, respiration in parallel to cellular proliferation and insulin production (basic and glucose-stimulated insulin secretion (GSIS)) in response to (I) AngIV (and its stable analogues) versus AngII, (II) pharmacological inhibitors of APA and APN, (III) genetic deficiency for APN, and (IV) the application of “stressors” including high glucose or direct application of H2O2. This will be analysed in isolated pancreatic islets from wild-type vs. APN-/- mice (already provided by Linda Shapiro [3]) and BRIN-BD11 rat insulinoma cells. Special focus will be given to redox-activated and redox-regulated signalling pathways that link ROS to the AngIV-IRAP/AT4R RAS axis and islet function. Essential collaborations include the analysis of CL composition and oxidation [Schild 2,4,5], mitochondrial membrane properties [B2 (Scholz)2,4,5], subcellular localisation and identification of relevant ROS sources [A4 (Lenzen/Elsner)] as well as relevant redox-regulated pathways and molecular targets [A1 (Lillig)].
Literature
1 Härdtner, C., Mörke, C., Walther, R., Wolke, C., Lendeckel, U.: Int J Mol Med. (2013) 32, 795.
2 Hermes, M., Scholz, F., Härdtner, C., Walther, R., Schild, L., Wolke, C., and Lendeckel, U.: Angew. Chemie (2011) 50, 6872
3 Winnicka, B., O’Conor, C., Schacke, W., Vernier, K., Grant, C.L., Fenteanny, F.H., Pereira, F.E., Liang, B., Kaur, A., Zhao, R., Montrose, D.C., Rosenberg, D.W., Aguila, H.L., Shapiro, L.H.: J Leukoc Biol. (2010) 88, 347.
4 Schild, L.*, Lendeckel, U.*, Gardemann, A., Wiswedel, I., Schmidt, C.A., Wolke, C., Walther, R., Grabarczyk, P., and Busemann, C.: Exp. Biol. Med. (2012) 237, 372. (*contributed equally)
Contact
Uwe Lendeckel
University Medicine Greifswald
Institute of Medical Biochemistry and Molecular Biology
Ferdinand-Sauerbruchstr.
D-17475 Greifswald
Tel: +49 (0)3834 86 5425
Fax:+49 (0)3834 86 5402
uwe.lendeckeluni-greifswaldde
Website